the evolution of lying: deception in the animal kingdom
Abstract
Deception is often regarded as a uniquely human moral failure. Yet misleading signaling is widespread across the animal kingdom. From mimicry and aggressive sexual signaling to tactical concealment in primates, deception emerges wherever communication affects fitness. This article synthesizes evidence from evolutionary biology, neuroscience and game theory to examine the mechanisms underlying deception. I distinguish deception from error, coercion, manipulation and self-deception, and evaluate whether lying destabilizes communication systems. Evidence suggests that deception is not a breakdown of communication, but an intrinsic and evolutionarily stable feature of signaling systems.
Introduction
Communication evolves because signals alter receiver behavior in ways that affect survival and reproduction. However, whenever signals influence fitness, natural selection also favors individuals capable of exploiting those signals. Deception therefore emerges as a structural feature of communication systems.
Biologically, deception refers to signaling that benefits the signaler while inducing a receiver response that would not occur under full information. Importantly, deception does not require conscious intent. It may arise from morphology, conditional behavior, reinforcement learning or higher-order social cognition.
Morphological Deception
Batesian Mimicry
The scarlet kingsnake (Lampropeltis elapsoides) closely resembles the venomous eastern coral snake (Micrurus fulvius). Both species exhibit red, black and yellow banding patterns, although their band order differs. Predators that have experienced or inherited avoidance of coral snakes often generalize this avoidance to kingsnakes.

The kingsnake does not possess venom. Its survival advantage arises from predator misclassification. This system evolves through negative frequency-dependent selection. When mimics remain rare relative to the venomous model, predators are more likely to avoid them. If mimics become too common, predators learn that the warning pattern is unreliable, reducing its protective value.
No cognitive awareness is required. The deception is encoded in morphology. However, the structure of deception is already present: signal, receiver inference and misalignment between signal and underlying state.
Aggressive Mimicry
Females of the genus Photuris provide one of the clearest examples of aggressive mimicry. These fireflies imitate the species-specific flash patterns of females from the genus Photinus. Male Photinus fireflies approach, expecting mating opportunities. Instead, they are captured and consumed.

The deception exploits a pre-existing sexual signaling system. Male Photinus rely on precise flash intervals to identify conspecific mates. Photuris females replicate these temporal patterns with high fidelity. The male’s perceptual system, tuned for mate detection, becomes the mechanism of his exploitation.
This form of deception does not require belief representation. It exploits sensory bias and signal recognition thresholds shaped by sexual selection.
Context-Dependent Behavioral Deception
Distraction Displays
The killdeer (Charadrius vociferus) is a ground-nesting bird whose eggs are vulnerable to terrestrial predators. When a predator approaches the nest, the adult bird performs a “broken-wing display.” It drags one wing along the ground, flutters erratically and vocalizes as though injured.

Predators are highly responsive to cues of vulnerability. The display redirects pursuit away from the nest and toward the apparently injured adult. Once the predator is sufficiently distant, the killdeer abruptly takes flight, revealing its intact condition.
This behavior is conditional and reversible. It is triggered by predator proximity and terminated when the threat diminishes. The deception exploits predictable predator pursuit strategies. However, there is no evidence that killdeer represent predator beliefs; the behavior may function as a fixed action pattern modulated by contextual input.
Strategic Cheating in Cooperative Systems
Cleaner Fish Mutualism
The bluestreak cleaner wrasse (Labroides dimidiatus) operates cleaning stations on coral reefs. Client fish visit to have ectoparasites removed. The interaction is mutualistic: clients gain parasite removal; cleaners gain nutrition.

However, cleaners prefer consuming client mucus, which is more nutritious than parasites. When cleaners bite mucus instead of parasites, clients often jolt or terminate the interaction. Some clients chase or avoid cleaners who repeatedly cheat.
Experimental evidence demonstrates that cleaners behave more cooperatively when potential clients are observing. This audience effect indicates that cleaners adjust behavior strategically in response to social context. The system resembles an iterated prisoner’s dilemma: cooperation and cheating coexist, constrained by punishment and partner choice.
Importantly, this system can be explained by reinforcement learning and repeated interaction dynamics without invoking full theory of mind.
Tactical Deception in Primates
Perspective-Taking in Chimpanzees
Studies of chimpanzees (Pan troglodytes) reveal sophisticated sensitivity to what conspecifics have or have not perceived. In experimental settings, subordinate individuals preferentially retrieve food that dominant individuals did not witness being hidden.
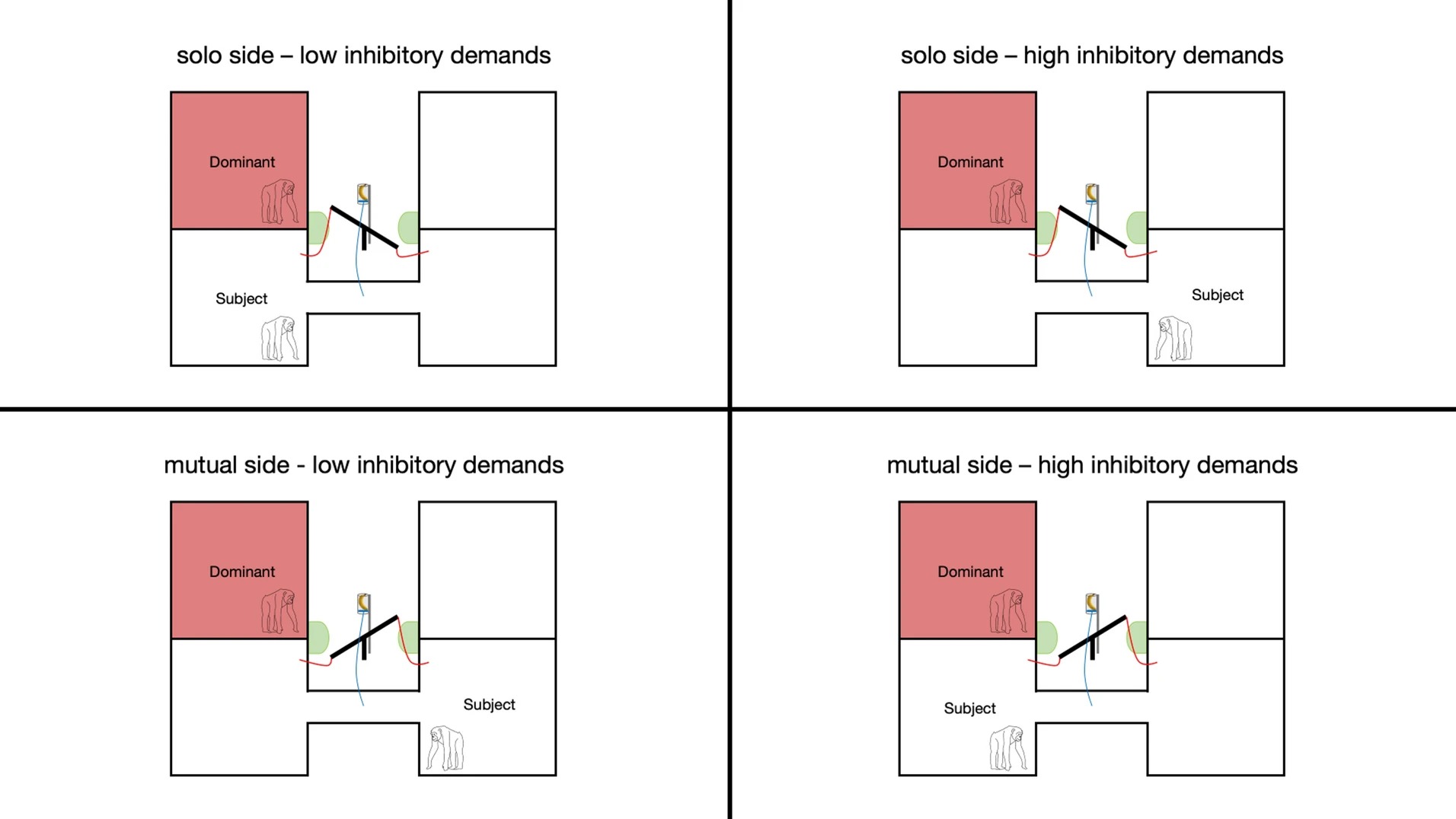
When given access to two food sources, one visible to a dominant competitor and one hidden, subordinates disproportionately approach the hidden resource. This suggests tracking of visual access and competitive knowledge asymmetries.
Chimpanzees also modify vocalizations depending on social context, suppressing food calls in competitive environments. These behaviors indicate perspective-taking at least at the level of perceptual access. Whether chimpanzees represent false beliefs remains debated, but they clearly track what others have seen.
Here deception approaches intentional concealment. It requires inhibitory control and social memory, capacities supported by expanded prefrontal cortices in primates.
Sexual and Resource Opportunism
Adélie penguins (Pygoscelis adeliae) construct nests from small stones that elevate eggs above melting ice. Stones are limited and highly contested. Observational reports describe extra-pair copulations occurring in exchange for access to stones.

While such exchanges occur, interpretation requires caution. There is no evidence that females signal long-term commitment and then deliberately violate it. A more parsimonious interpretation is opportunistic mating within a resource-constrained breeding colony.
This example highlights the importance of avoiding anthropomorphic interpretations of transactional mating behavior.
Evolutionary Game Theory
Evolutionarily Stable Strategies
A deceptive strategy is stable only if it yields higher fitness when rare and cannot be invaded by alternative strategies. Batesian mimicry satisfies this condition.
Handicap Principle
Costly signals may ensure honesty because only high-quality individuals can afford them. High production cost limits deceptive exploitation.
Iterated Prisoner’s Dilemma
Cleaner-client systems illustrate repeated-game dynamics. If cheating becomes too frequent, cooperation collapses. Thus deception remains constrained.
Game-theoretic models demonstrate that partial dishonesty can coexist with reliable communication. Systems collapse only when signal reliability falls below a threshold.
Neurobiological Foundations of Deception
Executive Control
In humans, deception recruits dorsolateral prefrontal cortex (working memory), ventrolateral prefrontal cortex (inhibition) and anterior cingulate cortex (conflict monitoring). Lying increases cognitive load relative to truth-telling.
Mentalizing Networks
Belief-based deception activates the temporoparietal junction and medial prefrontal cortex, regions associated with theory of mind. Nonhuman primates possess homologous but less specialized structures.
Reward Systems
Successful deception activates dopaminergic reward pathways. Reinforcement learning mechanisms likely stabilize deceptive strategies in many species.
Intentional lying requires representation of truth, representation of another’s belief, inhibitory control and alternative signal production. Clear evidence for full belief-based lying outside humans remains limited.
Conceptual Distinctions
Deception vs Error
Error is accidental misrepresentation. Deception is adaptive misrepresentation maintained by selection.
Deception vs Coercion
Coercion alters behavior through force or threat. Deception alters behavior through induced misclassification.
Deception vs Manipulation
All deception is manipulation, but not all manipulation is deception. Deception requires false signaling.
Self-Deception
In humans, self-deception may facilitate interpersonal deception by reducing cognitive dissonance. Evidence in nonhuman animals remains limited.
Is Lying Fundamentally Negative?
From an evolutionary perspective, deception is value-neutral. It is frequency-dependent, costly and constrained by detection mechanisms. If deception becomes too common, communication collapses.
Thus honesty and deception co-evolve. Deception is not a corruption of communication but an inevitable outcome of signaling systems shaped by natural selection.
References
- Abe, N. (2011). How the brain shapes deception. Neuroscientist.
- Bshary, R., & Grutter, A. (2002, 2006). Cleaner fish mutualism studies.
- Dunbar, R. (1998). The social brain hypothesis.
- Grafen, A. (1990). Biological signals as handicaps.
- Hare, B., Call, J., & Tomasello, M. (2001). Chimpanzee perspective-taking.
- Lloyd, J. (1965). Aggressive mimicry in fireflies.
- Maynard Smith, J. (1982). Evolution and the Theory of Games.
- Maynard Smith & Harper (2003). Animal Signals.
- Ritland & Brower (1991). Mimicry clarification.
- Ruxton et al. (2004). Avoiding Attack.
- Searcy & Nowicki (2005). The Evolution of Animal Communication.
- Spence et al. (2001). Neurobiology of deception.
- Trivers, R. (2011). The Folly of Fools.
- Zahavi, A. (1975). Handicap principle.